
纽迈生物超免技术依托全人源抗体转基因小鼠引领抗体研发新突破
在抗体发现领域 ,一直面临着一个棘手难题:当免疫原与小鼠的自体卵白高度相似时,会引发免疫耐受现象,即在B细胞和T细胞的成熟历程中,针对自体抗原发生的免疫细胞会被清除或抑制,无法获得强的免疫响应。例如肿瘤相关抗原GPC3(glypican-3)的人鼠同源性高达95%,炎症因子HMGB1人鼠同源性更是凌驾99%,用人的GPC3或HMGB1抗原免疫小鼠后,小鼠体内通常只能检测到极低滴度的抗体,甚至无明显免疫应答,这严重制约了针对此类靶点的抗体发现事情。
免疫耐受本质上是机体的一种掩护性机制,它能够制止机体发生针对自身靶点的免疫细胞或抗体,从而导致自身免疫疾病。然而,对于治疗性抗体的研发而言,免疫耐受却成了重大阻碍。它不仅限制了针对高同源靶点或低同源靶点的高同源区域的抗体发生,还淘汰了获得的抗体识别表位的多样性,使得抗体研发的难度大大增加。
为了攻克免疫耐受的问题,科学家们进行了诸多实验。
一种常见的战略是利用自身免疫病模型免疫。免疫耐受机制失衡将会引发自身免疫病,基于此原理,使用具有自身免疫病特征的动物模型进行免疫,可以增强免疫响应。但是,这类自身免疫病模型通常存在严重的自免表型,饲养难度大,繁育性能不佳。在经抗原刺激后,小鼠死亡率较高,这不仅降低了实验的乐成率,还大幅增加了实验成本。
另一种要领是敲除靶点基因。通过构建目标卵白缺失的小鼠,将抗原注射至此类小鼠体内,小鼠的免疫系统“首次”接触到该卵白,从而规避中枢耐受。对于部门靶点来说,这种要领具有一定可行性。然而,该要领需要针对每个靶点建设基因工程小鼠,成本高且周期长。更为要害的是,约1/3的小鼠卵白编码基因不能被敲除,因为这些基因缺失后小鼠会在胚胎期或出生后短时间内死亡。此外,像CD19, CD20等一些在重要免疫细胞上表达的基因,敲除之后会导致免疫细胞缺陷,使得小鼠无法用于免疫实验。
跨物种免疫也是一种实验。将人源或哺乳动物高度同源卵白注射至非哺乳动物(如鸡、龟或鲨鱼)体内,利用这些物种与哺乳动物亲缘关系远的特点,逃避对哺乳同源序列的耐受。例如,禽类(鸡)发生IgY抗体,鲨鱼能发生单域IgNAR抗体。这些物种往往能发生针对人源靶点的强免疫应答。不外,这种战略也存在显著缺陷,后续需要进行庞大且昂贵的工程革新和人源化处置惩罚。此外,非传统模型动物的免疫技术、单克隆化筛选和制备体系尚不成熟,难以形成高效工业化流程。
由此可见,现有的这些要领都难以彻底解决高同源目标诱导耐受的问题。基于此,纽迈生物建设了超免技术(HyperImmune Technology,HIT)。该技术通过对免疫耐受形成历程进行精准调治,抑制负筛选历程,从而实现在不敲除靶点基因的情况下,对高同源抗原发生强免疫响应,使高同源靶点、低同源靶点的高同源区域的抗体发现成为可能。即即是对于那些敲除后会导致小鼠致死,或在要害免疫细胞上表达的靶点,也能够高效、快速地获得高质量的抗体。
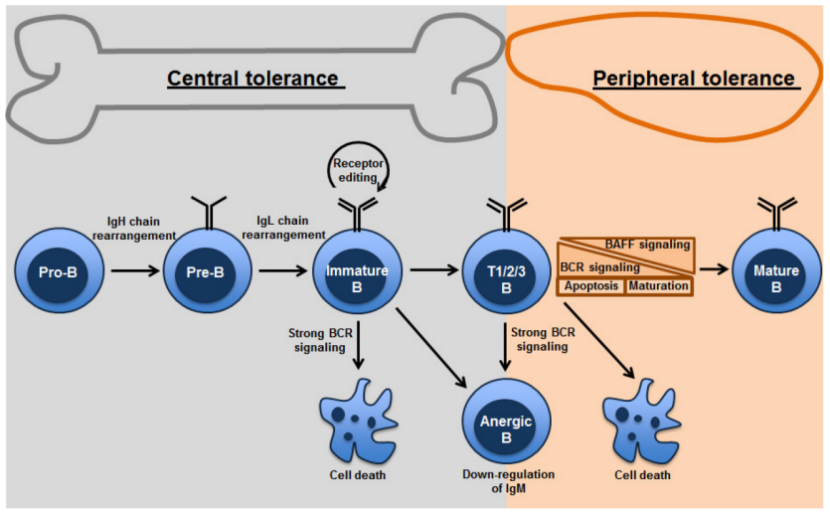
图一 B细胞的中枢耐受及外周耐受
超免技术(HIT)案例分享
靶点:HMGB1
人鼠同源性:99.1%
该靶点基因敲除后小鼠无法正常存活
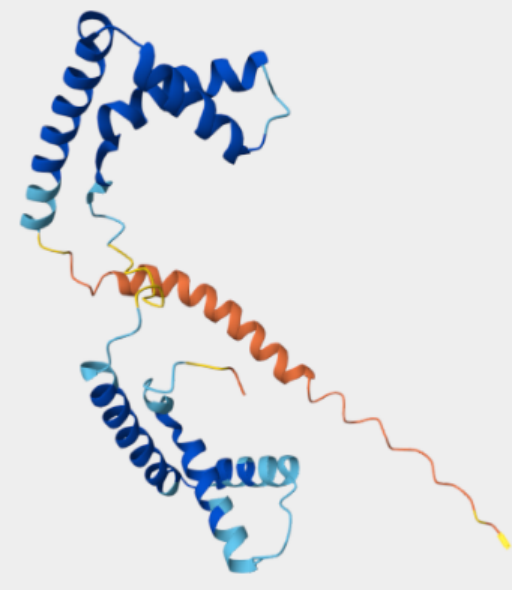
图二 人类HMGB1卵白结构

图三 人类HMGB1卵白功效域(人/鼠有2个氨基酸差异)
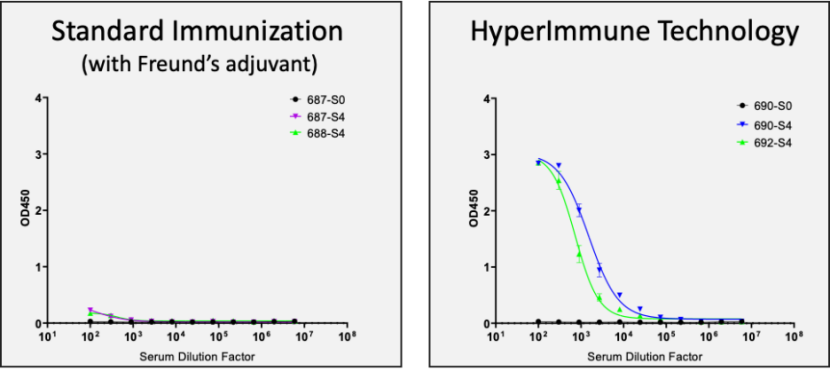
尺度免疫(弗氏佐剂)险些无响应;超免技术(HIT)响应良好
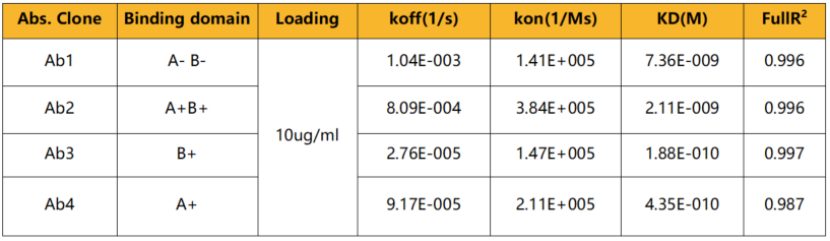
使用人类 HMGB1 卵白对 NeoMab 小鼠进行免疫实验,结果显示:接纳尺度免疫(弗氏佐剂)时,小鼠险些无免疫响应;而运用超免技术(HIT),小鼠则体现出良好的免疫响应。从超免小鼠中选择1只进行杂交瘤筛选,乐成获得了数百个特异性识别人类HMGB1卵白的克隆。经表位检测,获得了划分识别BOX-A、BOX-B、Acidic tail及其他区域的抗体分子,且这些克隆均能100%实现人鼠交织识别。挑选部门结合差异表位的分子进行亲和力检测,结果讲明其亲和力达可达10-9M——10-10M,这些抗体分子具备作为全人源治疗性抗体分子进行后续开发的潜力。
表一 全人HMGB1抗体亲和力(BLI检测)
关于纽迈生物
纽迈生物是尊龙团体生物的全资子公司。公司依托具有自主知识产权的全人源抗体转基因小鼠(NeoMab?-IgG小鼠、NeoMab?-CLC小鼠、NeoMab?-HC小鼠),专注于抗体开发服务和抗体分子的授权转让业务。致力于为客户提供高效、便捷的抗体药物发现技术服务,立志成为抗体药物发现领域值得信赖的相助同伴。
联系我们:BD@neomab-bio.com
参考文献
1. Cerottini, J. C., P. H. Lambert and F. J. Dixon (1969). "Comparison of the immune responsiveness of NZB and NZB X NZW F1 hybrid mice with that of other strains of mice." J Exp Med 130(5): 1093-1105.
2. Chen, R., R. Kang and D. Tang (2022). "The mechanism of HMGB1 secretion and release." Exp Mol Med 54(2): 91-102.
3. Gururajan, M., V. J. Sindhava and S. Bondada (2014). "B Cell Tolerance in Health and Disease." Antibodies 3(1): 116-129.
4. Jiang, X., L. Sun, C. Hu, F. Zheng, Z. Lyu and J. Shao (2023). "Shark IgNAR: The Next Broad Application Antibody in Clinical Diagnoses and Tumor Therapies?" Mar Drugs 21(9).
5. Liptak, N., Z. Gal, B. Biro, L. Hiripi and O. I. Hoffmann (2021). "Rescuing lethal phenotypes induced by disruption of genes in mice: a review of novel strategies." Physiol Res 70(1): 3-12.
6. Ofuji, K., K. Saito, T. Yoshikawa and T. Nakatsura (2014). "Critical analysis of the potential of targeting GPC3 in hepatocellular carcinoma." J Hepatocell Carcinoma 1: 35-42.
7. Percival-Alwyn, J. L., E. England, B. Kemp, L. Rapley, N. H. Davis, G. R. McCarthy, J. B. Majithiya, D. J. Corkill, S. Welsted, K. Minton, E. S. Cohen, M. J. Robinson, C. Dobson, T. C. Wilkinson, T. J. Vaughan, M. A. Groves and N. J. Tigue (2015). "Generation of potent mouse monoclonal antibodies to self-proteins using T-cell epitope "tags"." MAbs 7(1): 129-137.
8. St Clair, E. W., D. Kenan, J. A. Burch, Jr, J. D. Keene and D. S. Pisetsky (1991). "Anti-La antibody production by MRL-1pr/1pr mice. Analysis of fine specificity." The Journal of Immunology 146(6): 1885-1892.